


纳米抗体(Nanobodies,HCAb,Nb,sdAb,VHH)简介
骆驼科动物的血清同时含有常规的异四聚体抗体和独特的功能性重(H)链抗体(HCAb)。这些同二聚体抗体的H链由一个抗原结合结构域,纳米抗体(VHH)和两个恒定结构域(CH2,CH3区)组成。由于第一恒定结构域的缺失和纳米抗体(VHH)侧的重塑表面(通常与常规抗体中的L链相关),HCAb不能掺入轻(L)链。已经鉴定了构成HCAb的遗传元件,但是仍在很大程度上对将这些抗体从其专用基因体内生成为抗原特异性和亲和力成熟的真正抗体进行研究。然而,由多个晶体结构支持的抗原特异性VHH的简便鉴定及其有益的生化和经济特性(大小,亲和力,特异性,稳定性,生产成本)鼓励了将这些单域抗体用作研究工具以及生物技术和医学的抗体工程化。
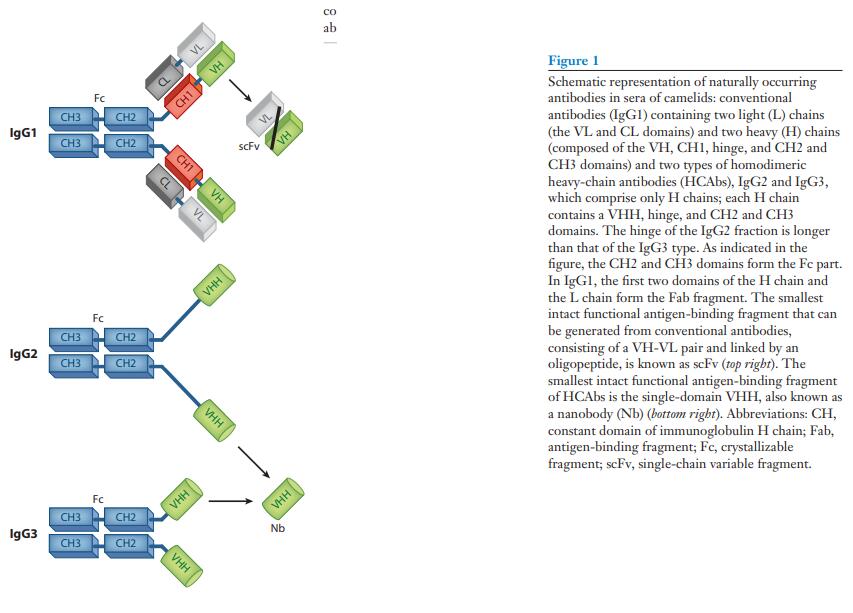
由两条相同的重(H)链和两条相同的轻(L)链多肽组装而成的免疫球蛋白-γ(IgG)抗体的整体结构已在哺乳动物中得到了很好的确立和高度保守(图1)(1)。 这些免疫球蛋白的L链包含两个结构域,而H链折叠成四个结构域。 H和L多肽链的N末端结构域的序列在抗体之间变化(称为可变结构域,即VH和VL)。 配对的VH-VL结构域构成识别抗原的可变片段(Fv)。 其余的H和L序列更为保守(分别缩写为CH和CL)。 最后两个CH区域对于募集免疫细胞(例如巨噬细胞和天然杀伤细胞)或效应子功能(例如补体激活)很重要。
![]()
纳米抗体(nanobody,Nb), 即重链单域抗体VHH (variable domain of heavy chain of heavy-chainantibody)
在骆驼科(2)的血清中发现了这种常规哺乳动物IgG结构的一个典型的例外。 除了常规的异四聚体抗体外,这些血清还具有特殊的IgG抗体。 IgG抗体被称为重链抗体(HCAb),不含L链多肽,并且由于缺乏第一个恒定域(CH1)而具有独特性(图1)。 同型二聚体蛋白的H链在其N端区域包含一个专用可变域,称为纳米抗体(VHH),用于与其同源抗原缔合。 HCAb中的VHH与常规抗体的Fab片段(抗原结合片段)的结构和功能等效(图1)。
骆驼科生物家族包括:
骆驼科是亚足类亚目( suborder Tylopoda)中唯一的现存家族,与反刍动物(Ruminantia )(牛,山羊,绵羊,羚羊及其他)和Soiformes(猪和河马科)一起构成木偶目( Artiodactyla)。尽管所有骆驼科动物的血清中均含有HCAb,但其他偶蹄动物(如Suiformes和Ruminantia)却没有这种功能性HCAb。然而,据报道,由VH和CH1区域的重要部分的基因缺失导致的HCAbs发生在患有病理性疾病的人类血清中(3,4)或在小鼠杂交瘤中(5)。由于截短的VH区和VL的缺失,这些HCAb在抗原结合中不起作用。引人注目的是,缺乏L链且缺乏常规CH1的免疫球蛋白也出现在哺乳鲨,沃伯贡(wobbegong)和可能发现的斑尾鱼( spotted ratfish)中(6)。这些Ig-NAR祖先抗体具有可变区,称为V-NAR,用于抗原识别。尽管Ig-NAR和骆驼HCAb的可变(V)序列非常不同,但它们显示出令人惊讶的结构和功能趋同进化(7)。
HCAb 的抗原结合片段包含一个单一可变结构域,可在对骆驼科动物进行免疫,克隆血液中循环的B细胞的VHH组成以及通过噬菌体展示淘选后,直接鉴定抗原结合的纳米抗体(VHH)(8)。 尺寸在纳米范围内的重组抗原特异性单结构域VHH也称为纳米抗体(Nb)或单结构域抗体(sdAb)。 在这篇综述中,我们总结了HCAbs和VHH结构域的主要结构特征,并提供了骆驼科中HCAbs可能产生机制的更新。 最后,我们回顾了Nbs的生化特性,这些Nbs构成了研究,生物技术和医学中各种常规应用和创新应用的基础。 铁汉 18:16:53
IgG中L链的缺失和H链中CH1的缺失是骆驼类HCAb的关键特征。 因此,其较小的分子重量为90,000,而不是常规抗体的150,000,而且其结构更为紧凑,可能更适合进入隐蔽靶标。 相反,尽管有人推测包含Pro-Gln重复序列的长IgG2长链可能形成结构上取代CH1区的延伸间隔子,但一个HCAb中两个互补位之间的较短距离可能会损害其交联抗原的能力。 (图1)
HCAbs和常规IgG在骆驼科动物血清中的百分比是可变的:在骆驼(camels)中,它可能达到50–80%,而在南美骆驼科动物(South American camelid species)中,它总计高达10–25%(9)。如此高的比例证明了HCAb在骆驼科动物的免疫保护中的重要性。
骆驼科动物的血液中循环着几种没有L链的IgG亚型,尽管由于将亚型纯化为同质性以及将亚型与同种型区分开来的困难,确切的数量仍存在争议。然而,同二聚体和异四聚体抗体很容易通过蛋白质G和蛋白质A的差分亲和色谱分离,其中常规抗体在较低的pH值下从蛋白质G柱上洗脱(2,10)。未保留在蛋白G上的单峰IgG组分被吸附在蛋白A上,仔细的pH控制洗脱可得到两个不同的HCAb-IgG2组分,分别为IgG2a和IgG2b(10)。研究人员表明,这些吸附剂的馏分可能仍含有几种亚型。针对不同蛋白A / G组分的IgG产生的单克隆抗体用于筛选美洲驼血清免疫球蛋白,证实了常规IgG的两种同种型以及IgG2和IgG3组分中分别存在三种和两种亚型(11,12, 13)。
对单峰骆驼B淋巴细胞的IgG cDNA的分析揭示了常规抗体存在两种IgG亚型,而HCAbs包含多种IgG亚型(14)。 各种cDNA序列仍需要与从蛋白A和蛋白G色谱或抗同型单克隆抗体获得的亚型(或同种型)匹配。 尽管不确定骆驼科动物血液中循环的IgG亚型的确切数目,但该数目仍在其他artiodactyls的IgG范围内。 已从牛中鉴定出三种IgG同种型(15),而猪含有无法通过蛋白A和/或蛋白G色谱法分离的六种IgG同种型(16)。
人类和小鼠抗体的研究表明,IgG亚型之间存在明显的分工。 尽管还尚不清楚各种骆驼科动物IgG亚型的确切作用和功能的知识,但受感染或接种疫苗的动物会在这三种同种型组分中引起免疫反应(不同动物在不同程度上,取决于实际的免疫原)。 值得注意的是,美洲驼IgG1和IgG3中和了西尼罗河病毒,而IgG2在这方面似乎不太有效(17)。 但是,同一项研究表明,来自所有三种同种型的IgG均与单核细胞和巨噬细胞的表面结合,表明它们能够募集免疫细胞。
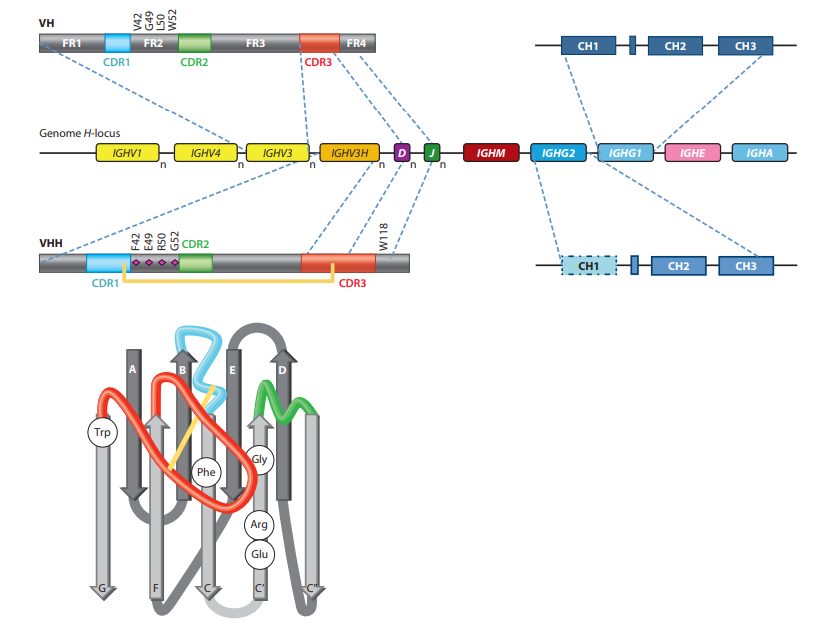
V域内的序列变异性位于三个更易变的框架(FR)区域包围的三个高变(HV)区域中(图2)(1)。折叠的V结构域包含九个β链(ABCC'-C” -DEFG),它们以四链β-折叠和五链β-折叠组织,通过环和通过Cys23和Cys94之间的保守二硫键连接,与保守的Trp相对。在此体系结构中,HV区位于分别连接BC,C'-C'‘和FG链的环H1至H3中,并且聚簇在域的N末端,形成连续表面与表位表面互补,因此命名为互补决定区(CDR)。尽管loop中的顺序为HV,但长度变化不受H3 loop的限制。令人惊讶的是,H1和H2 loop中的Cα位置占据限制性位置。显然,在人或小鼠的VH结构中仅观察到少数所谓的规范环结构,甚至loop结构组合的数量也受到限制(18)。因此,loop长度和loop中关键残基的存在可预测其实际架构。在常规抗体中,VH的三个HV环与VL的三个HV环并置,并提供约600–900?2的平台(确切的相互作用表面取决于环的大小及其氨基酸,如以及用于计算的算法)。总体而言,常规抗体的互补位形成空腔,凹槽或平坦表面(突出了单个氨基酸侧链的微小起伏),这些结构已与识别小分子,线性肽和较大的抗原( (例如蛋白质)(19)。
![]()